Archives
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-04
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-11
- 2018-10
- 2018-07
-
It is well established that Shp can function as
2020-07-07
It is well established that Shp-2 can function as a substrate for several RTK, such as PDGF or EGFR [23], [36]. To test whether DDR1 recognizes Shp-2 as its substrate, we overexpressed a catalytically inactive Shp-2 mutant together with DDR1b in 293 cells [31]. Immunoprecipitation of Shp-2 followed
-
br Acknowledgements br This work was supported by grants
2020-07-07
Acknowledgements This work was supported by grants from the National Natural Science Foundation of China (Grant No.: 30260088) and the prophase research for National “973” Project of China (Grant No.: 2007CB116203). The DNA sequences of Hbvp1 and the Hbvp1 promoter were deposited in GenBank, an
-
In a study performed simultaneously by another group
2020-07-06
In a study performed simultaneously by another group, EWS-FLI-1 introduction into unsorted murine bone marrow-derived cytidine resulted in tumors with various phenotypes including one that is consistent with that of Ewing sarcoma [68]. The tumors expressed markers associated with Ewing sarcoma and
-
Among the Ub interactions observed
2020-07-06
Among the ~Ub interactions observed in the HOIP structure is one that involves a RING2 domain (from the non-cognate HOIP) [47]. A similar binding interaction was defined between HHARI RING2 and the hydrophobic patch on Ub by NMR [54]. HHARI RING2 mutations that ablate the interaction inhibit formati
-
Studies of Jelski and Szmitkowski show
2020-07-06
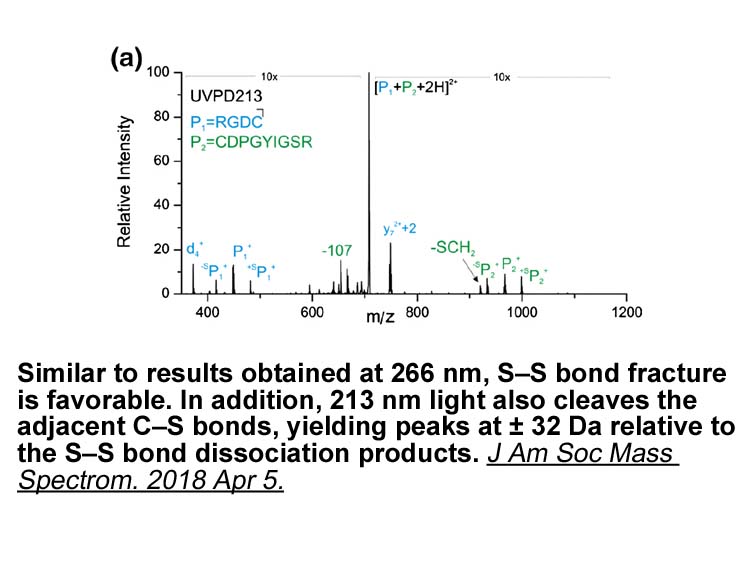
Studies of Jelski and Szmitkowski show that differences of alcohol dehydrogenase (ADH) and aldehyde dehydrogenase (ALDH) activity between U-104 synthesis of many cancers and healthy tissue may be one of the factors intensifying carcinogenesis [7]. Moreover, changes of ADH and ALDH activity in cancer
-
br Discussion The differences in the kinetic properties
2020-07-06
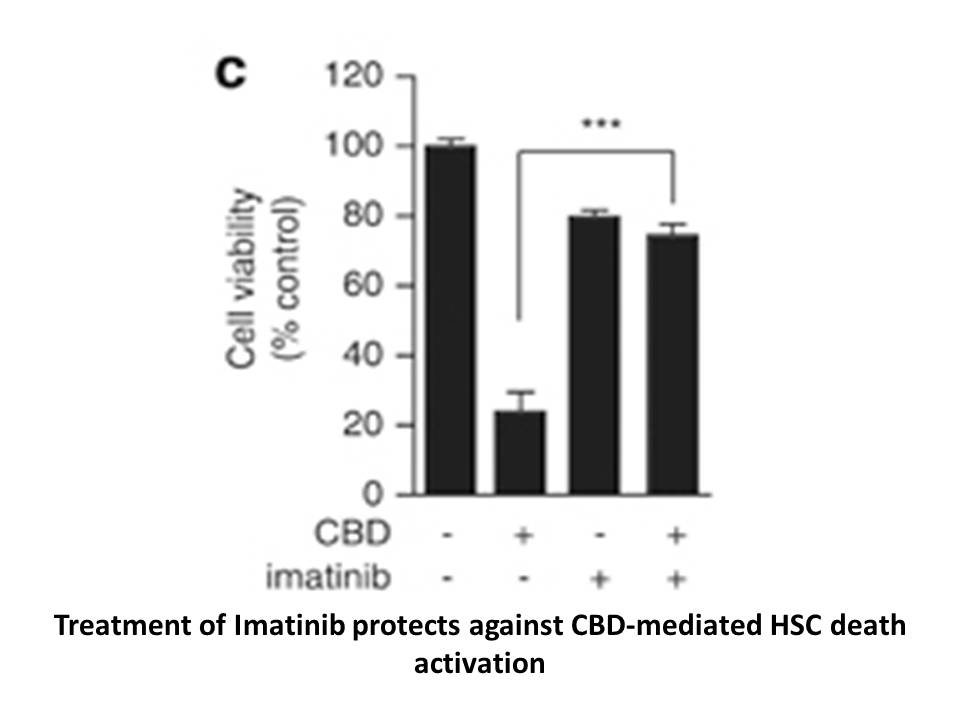
Discussion The differences in the kinetic properties of L-AmDH and LeuDH presented herein reflect a different kinetic mechanism. When compared to its parent enzyme, L-AmDH displayed a different substrate binding order, lower affinity for the keto substrate, and stronger product inhibition. The tw
-
br Materials and methods br Results The BUN
2020-07-06
Materials and methods Results The BUN and creatinine levels in the saline-treated CRF group were found to be significantly higher than those in the sham-operated control rats treated with either montelukast or saline (ppla2 inhibitor tissues, were found to be significantly higher in the saline
-
Cells expressing the CDK S D mutant
2020-07-06
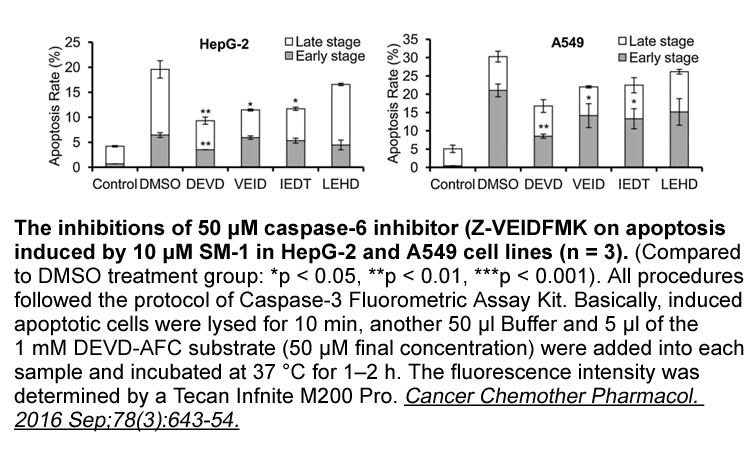
Cells expressing the CDK5 S47D mutant exhibit impaired migration and enhanced proliferation: Finally, we wished to determine the functional significance of this phosphoevent. Multiple studies have implicated active CDK5 in promotion of cell migration [9,16,[21], [22], [23], [24]] and in one study th
-
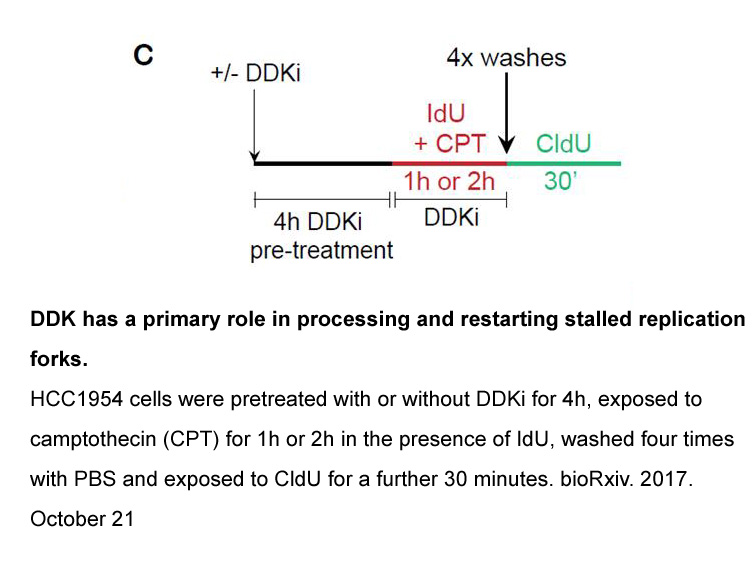
The paper is organized as follows http www apexbt com
2020-07-06
The paper is organized as follows. Section 2 gives a brief presentation of the JPEG-LS algorithm and respective hardware implementation. In Section 3 the fault injection model and the simulation-based susceptibility analysis method are reviewed. Section 4 provides a discussion on the susceptibility
-
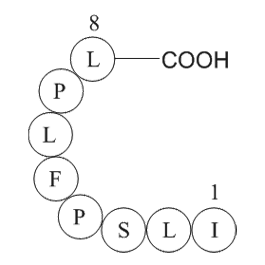
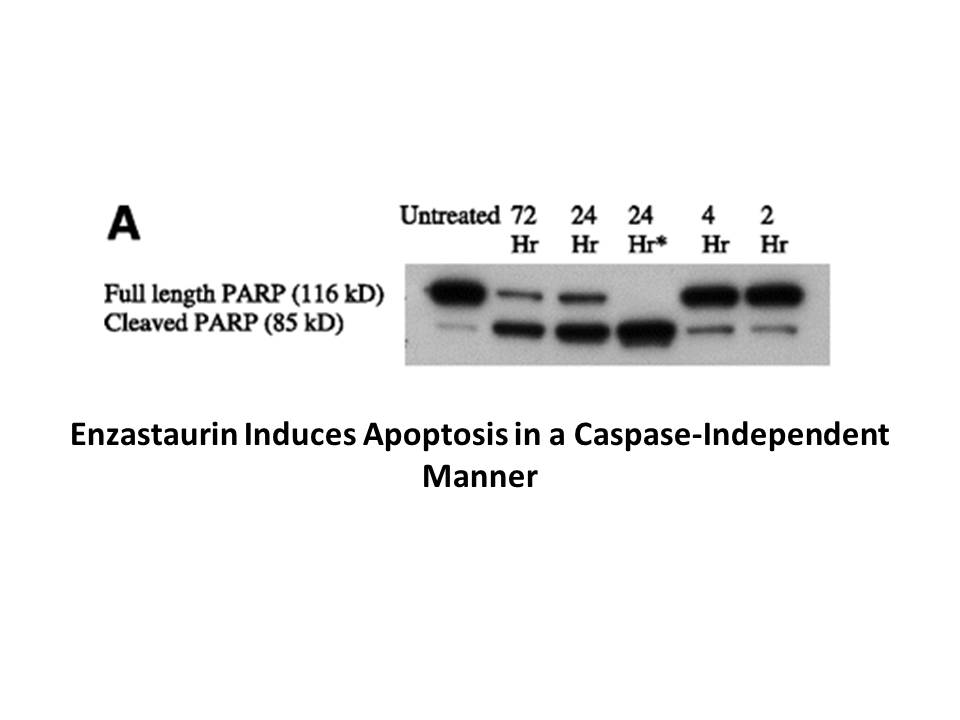
Keratin K has been identified as the
2020-07-03
Keratin 8 (K8) has been identified as the first intermediate filament expressed during development in amphibians and mammals. In zebrafish, a type II cytokeratin 8 (zf-K8) cDNA has been cloned. zf-K8 gene was maternally inherited and expressed in all surface cells throughout the embryonic stages old
-
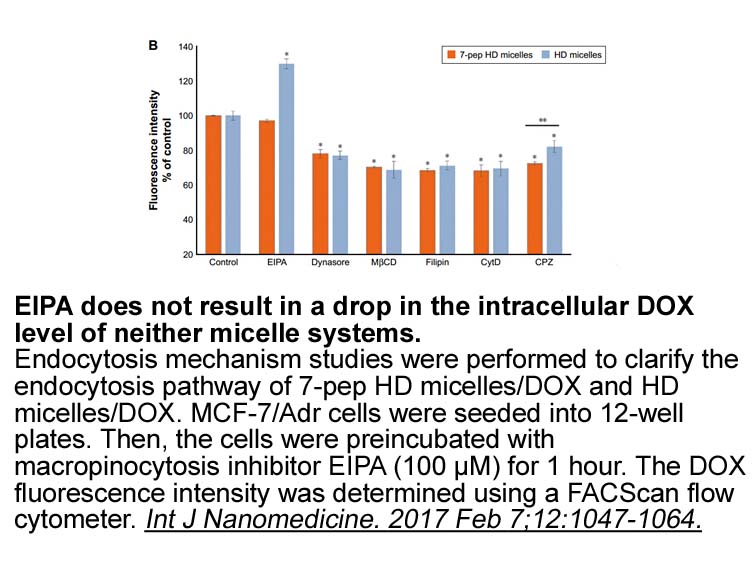
br Materials and methods br Statistical
2020-07-03
Materials and methods Statistical analysis All statistical analyses were performed using SPSS software version 20. Protein lxr agonist data of DNMTs and clinicopathological parameters were collected. Mann-Whitney U Test was used to assess the differential expression of nonparametric data of D
-
In order to derive a
2020-07-03
In order to derive a better understanding of the results from our SAR study, we utilized Glide to model the binding of the SAR compounds with the non-active site pocket of TS-DHFR shown in C. The majority of models position the phenolic moiety of these compounds within the non-active site pocket, wh
-
To better understand Wolbachia s dependence on
2020-07-03
To better understand Wolbachia’s dependence on these enzymes, RNAi experiments targeting these transcripts will be required to examine the consequences of their down-regulation on Wolbachia in the different tissues of the parasite. Initial studies have shown that decreases in Bm-cpl-3 and Bm-cpl-6 t
-
Materials and methods Construction of the
2020-07-03
Materials and methods Construction of the Chk-YB-1b-Neo vector. Approximately 107 plaques from a chicken genomic library established in EMBL3 vector (Clontech Palo Alto, CA) was screened and one clone containing a 12kb insert was selected. Digestion of this clone with SalI and EcoRI restriction enz
-
TMC647055 Choline salt sale A limitation of this study is th
2020-07-03
A limitation of this study is the between-study heterogeneity of the datasets analyzed. To address this heterogeneity, we used a random-effects model in our analysis. Also, only 2 of the databases provided information on sex, and as a result, the sex-specific analyses were performed with a smaller c
16264 records 821/1085 page Previous Next First page 上5页 821822823824825 下5页 Last page